Biosemiotics and the Origins of Life on Earth (Part 8 of 8)
0316 Oxygen gas is a byproduct of photosynthesis. Over billions of years, the continual release of oxygen transforms the atmosphere of the Earth.
The ubiquity of oxygen gas in today’s atmosphere makes experimental research into the chemistry of the early Earthdifficult. Today, the reaction that Sharov suggests, the oxidation of an alkane to a fatty acid, would require elaborate precautions. Why? Even a trace amount of oxygen would directly react with the light-absorbing pigment.
0317 So, what am I saying?
Well, research is difficult.
0318 Also, as soon as one gets to the earliest forms of life on Earth, such as photosynthetic prokaryotes, the “genomic complexity” (nominally, the length of DNA that belongs to only functional genes) is already high. If one plots the genomic complexity of (1) prokaryotes, such as bacteria, (2) single-celled eukaryotes, such as amoebas, (3) multicellular water animals, such as fish (4) invertebrate land animals, such as worms, and (5) vertebrate land animals, such as mammals, versus time for first fossil evidence, one gets the following graph.
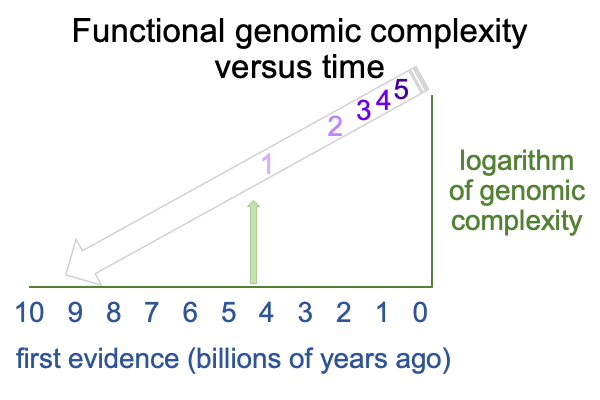
0319 On one hand, Sharov concludes that the genomic complexity doubles every 340 million years since the start of the Earth.
On the other hand, Sharov points out that, if one projects the line down to zero genomic complexity, the intersection occurs a little over 9 billion years ago. But, the Earth is only 4.5Byr.
Fortunately, the universe is around 15 billion years old.
0320 If the early Earth is seeded, then biologists already have a label, “panspermia”.
All other planets and moons in the solar system should be similarly seeded.
So, future space exploration may provide an answer.
If it turns out that the early Earth is seeded through panspermia, then research into the origins of life (in general) becomes even more difficult.
0321 Now, I conclude.
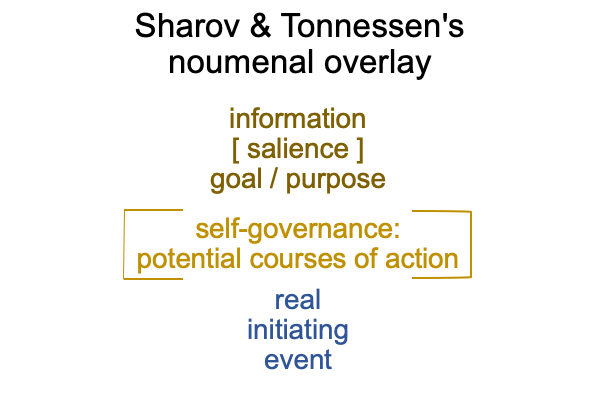
Sharov and Tonnessen’s noumenal overlay characterizes biosemiotics.
The Deacon-Tabaczek interscope characterizes emergence.
Both relational structures apply to inquiry into the origin of life on Earth.
This examination demonstrates how the two relational structures relate to one another and constitute complementary approaches for further inquiries into the origins of life.
0322 But, what I have learned concerns more than the topic of the origin of life.
This is significant.
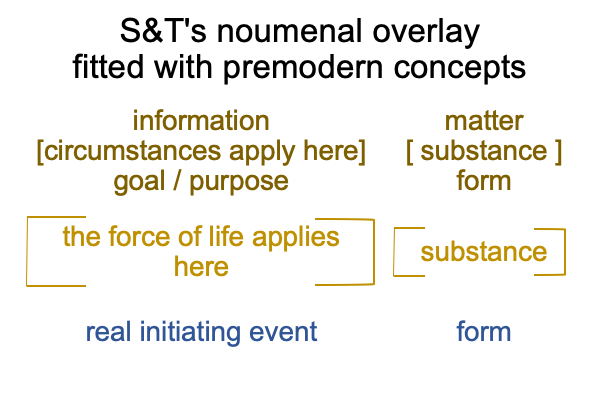
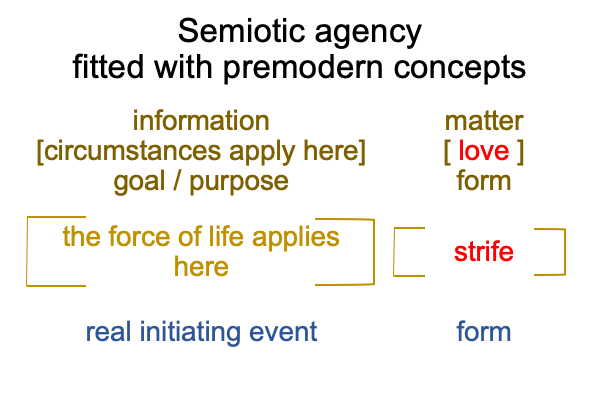
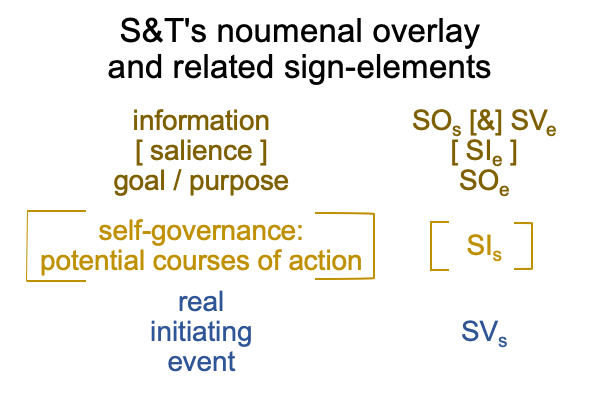
Sharov and Tonnessen’s noumenal overlay may “expand” to include the entire D-T interscope, which includes both the specifying and the exemplar sign-relations.
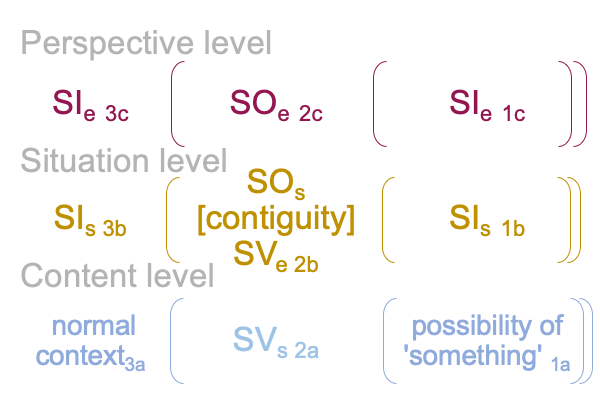
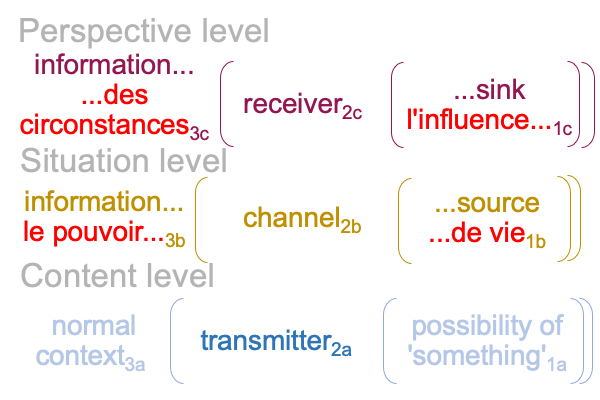
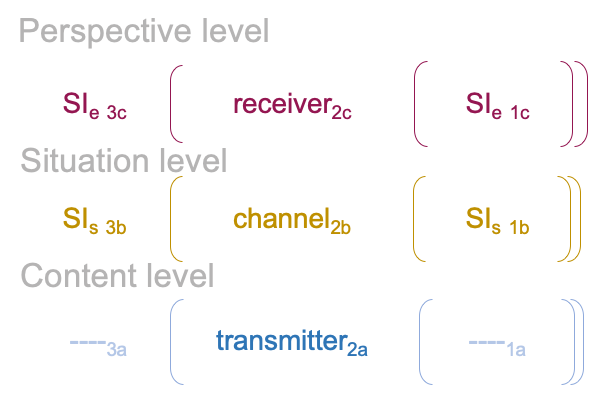
0322 By extension, the S&T noumenal overlay associates to any three-level interscope, containing two sign-relations,according to the comparison in the following figure.
0323 The topic of the origin of life on Earth turns into a valuable insight into biosemiotics, emergence, and two sign-relations.