0299 Here is a three-level interscope for emergence, with the specifying and exemplar sign-relations noted.
0300 Emergence enters into S&T’s noumenal overlay.
Here are the results.
0301 When Sharov and Tonnessen confront the origin of life on Earth in chapter five of Semiotic Agency, associations with Mariusz Tabaczek’s formulation of emergence are not apparent. The focus on inquiry is on chemical self-replication rather than structures that capture thermodynamic energy2a through an emergent2b then dissipate the emergent’s energy2cby building a persistent structure.
The eukaryotic cell’s metabolism of glucose and oxygen works by extracting energy released in the combustion of these reagents.
In combustion, oxygen gas directly takes electrons from glucose, without any homeodynamics. Covalent bonds are broken. Covalent bonds form. Lots of free-energy is released and converted into heat.
In the eukaryotic cell, electrons produced by the oxidation of glucose (at one cellular location) are used to produce an emergent being, such as ATP, before going into the reduction of oxygen (at another cellular location), where more ATP is produced. ATP2b is the emergent being, whose energy is dissipated on the morphodynamic level.
0302 So, where is a scientist supposed to start, when considering abiogenesis?
Researchers into the origins of life focus on the formation of covalent bonds that constitute polymers. Polymeric molecules are persistent structures. But, scientists have not identified an emergent, similar to ATP, whose concentration is low yet constant, because it is produced on the homeodynamic level and used up on the morphodynamic level. Nor have researchers identified any thermodynamic processes amenable to exploitation by a homeodynamic level.
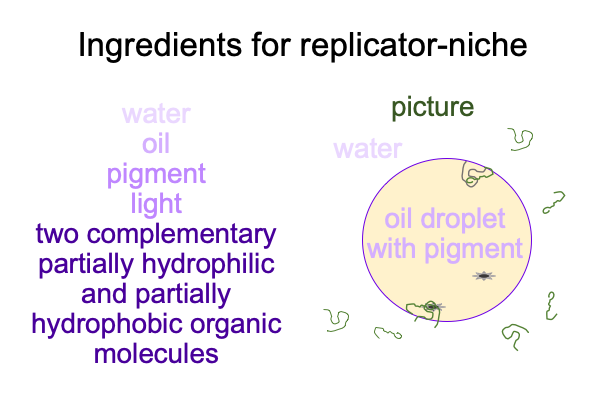
0302 In section 5.5 of Semiotic Agency and sections 9.4 and 9.5 of Pathways, Alexei Sharov presents a replicator-niche coupling model. Several items are required: water, oil, pigments (in oil), light, and two complementary molecules that are separate in water, yet combine to form an active site when they attach to the surface of an oil droplet.
0304 Here is a picture.
0305 Let me start with the oil droplet.
Water tends to drive alkanes out of solution. That is why alkanes form oil droplets in water. These droplets are not really stable, because they are not held together because of mutual attraction, but are held in place by the fact that each water molecule networks with other water molecules so well that, if a molecule does not participate in water’s hydrogen-bond networks, it gets driven out of solution. That also applies to the pigment, which is oil-soluble and not water-soluble.
0306 What about the polymers?
Parts of complementary polymers are soluble in water. Other parts are not as soluble. So, parts are driven out of water and parts are pulled back into water. These molecules collect on the surface of the oil droplet, then couple with one another, with the pigment and with an alkane, which is part of the oil droplet.
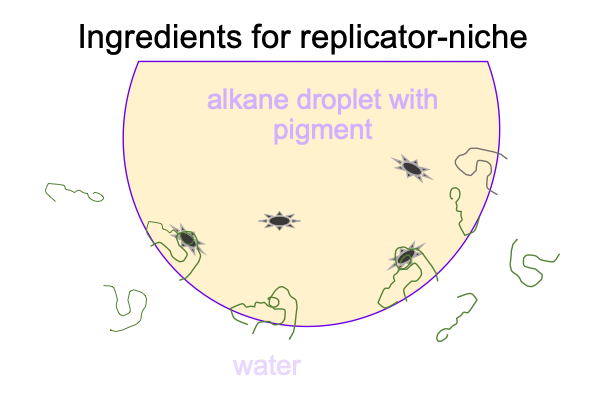
0307 There are no oxygen molecules in the picture. Today, the Earth’s atmosphere is around 20% oxygen and 80% nitrogen. In the early Earth’s atmosphere, reduced carbon compounds make the smaller fraction and nitrogen makes the large fraction. More or less. No scientist can go back in time and measure the composition of the atmosphere of the early Earth.
Reduced carbon in the atmosphere goes with the alkanes in the oil. Much of the light of the early sun is absorbed by the carbon-rich atmosphere, but some makes it down to pigments in the oil droplet. The pigment and complementary polymers conjoin in two locations in the figure below.
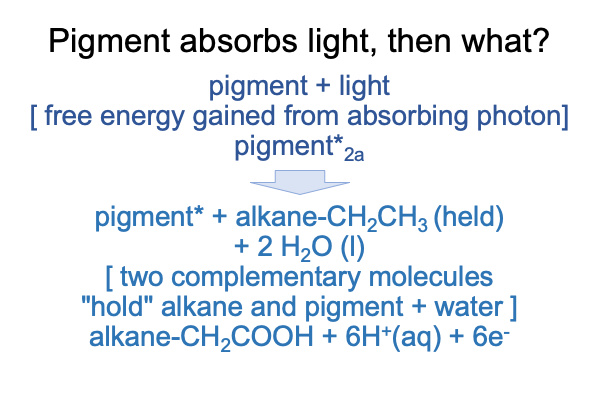
0308 Then, what happens?
The pigment absorbs a photon and becomes an electronically excited pigment.
Then, the energy captured by this pigment initiates a chemical reaction, where the alkane is oxidized to a fatty acid. Oxidation releases electrons. One among many possible oxidations is pictured above. With a little more oomph, that carboxylic acid would pop off as carbon dioxide. However, this reaction stops as an alkane chain with a carboxylic acid at the terminus. I call this molecule a “fatty acid”.
In the following figure, the two processes are depicted as two dyads. Each dyad exhibits the structure of reagents [turn into] products.
0309 Now, theoretically a reduction reaction is close at hand. If the oil droplet is near a chemical that can accept the electrons, then a coordinated reduction can take place. For example, the hydrogen ions and the electrons can combine to form 3H2(g). Or, atmosphere nitrogen (N2) can be reduced to ammonia, 2NH3.
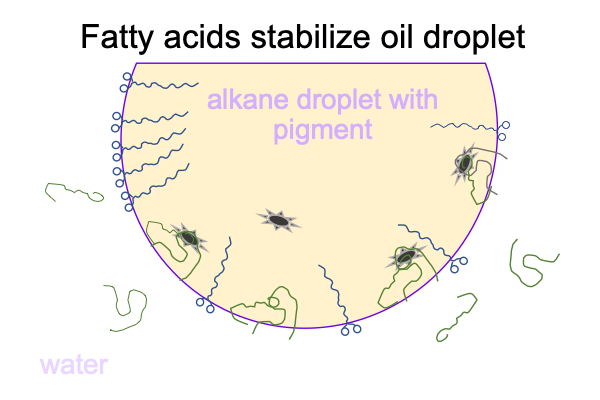
The fatty acid serves as the emergent being2b, because the carboxylic-acid side tends to favor the water and the alkane side stays in the oil droplet.
0311 In short, fatty acid is the emergent being2b that has the potential of stabilizing oil droplets1c, allowing them to “feed off” or “absorb” oil from less stable oil droplets2c.
0312 Does Sharov’s scenario, as far as it goes, fit the Deacon-Tabaczek interscope?
Indeed, it does.
0313 Does this interscope associate to the S&T noumenal overlay?
Yes, it does.
0314 Well, so far so good.
Nevertheless, there is a long way to go to get to a prokaryotic cell (as noted in sections 5.8-5.10 of Semiotic Agency and 9.6-9.8 of Pathways).
For example, prokaryotic cells replicate themselves through cell division. But, the replication is nothing like this oil droplet example. That is because DNA plays a role in prokaryotic cell division. Biologist call this type of replication, “template based”.
Also, there is the issue of the cell membrane. The cell membrane is a lipid bilayer, consisting of phosphorylated fatty acids. In other words, fatty acids may stabilize an oil droplet. Once those fatty acids have a phosphate attached to them, then their phosphates love water so much that the alkane-portion of the molecule is excluded from the water so strongly that a bilayer is stable.
0315 Also, there is the formalization of pigments that capture sunlight in order to produce energy-rich sugar molecules. Today, photosynthesis absorbs carbon dioxide (gas) and releases oxygen (gas). In the early Earth, photosynthesis does the same.
0333 Well, what if matter… er… information… consists of multiple biomolecules that are… um… modular… in so far as they… because of circumstances… simultaneously engage in a cooperative endeavor… a “love”, so to speak?
Some would say that this what if is similar to the hierarchical relation between parts and a whole.
But, would the whole be the form associated with goal or the form associated with the real initiating event?
Or, would it be the thing called “semiotic agency”?
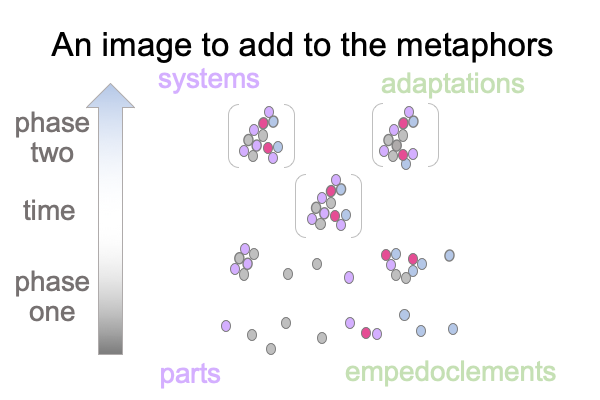
0334 At the start of section 10.2, the author lays this ambiguous multiple metaphor onto the procrustean bed of an evolutionary paradigm, where (on a molecular level) biological parts are added to one another in a piecemeal way to an evolving system.
The author proposes a phylogenomic-based biphasic model of module creation that explains evolutionary growth in biochemical systems. In phase one, modules nest within one another, in a provisional sort of way, until over time or suddenly, the form that is goal clarifies. Different modules start to work together as a semiotic agent. In phase two, modules working within that functioning cooperative change as semiotic agency diversifies.
0335 The second phase associates to “adaptation”.
The first phase does not have a proper name. I suggest the term, “empedoclement”.
An empedoclement is the inverse of an impediment.
0336 Here is an analogy.
Recently, I joined a tennis club. I trained to play the game by enduring real initiating events and reviewing information on my performance. Since I am so modular, different aspects of me perform independently of one another, so I did not adapt well to the circumstances. My instructor says that if I don’t think about what I am doing with each of my modules,then I could move holistically in a coordinated manner. I will become an adept, rather than a lackluster player full of impediments.
0337 One would think that routinely training and playing tennis would lead to (perhaps slow, but) steady improvement towards the goal of being competitive. This is a matter of adaptation. All the facets of mind and body are modular. Each module develops along its own trajectory. In apparent stasis, improvement is held at bay by one or two recalcitrant modules. In surprisingly fast change, two or more modules improve in tandem. The fast change could be an empedoclement.
0338 While this analogy helps, I find it difficult to imagine that first phase, where parts are present and may afford some advantages, but no one part realizes the game that is afoot.
0339 The author offers two images as metaphors for the phylogenomic-based biphasic model.
One metaphor is a tree, where the roots represent modules, the trunk stands for the modules coalescing into matter [love] form, where matter is information and the form is a telos or an end.
The other metaphor portrays modules themselves, which over time, interact to generate scaffolds and active sites, until a robust combination constellates.
Here is a picture of the second metaphor.
0340 The word, “constellates”, is a psychological term (actually, Jungian) that marks the coming into presence (esse_ce) of an archetypal form (essence). The result may be called “a primordial image”.
For example, the archetype of the king may constellate in a variety of ways, including the duplicitous and the honest, the greedy and the beneficent, as well as the foolish and the wise. Each pair of these primordial images informs us of a module within the one archetype and how that module can yield different responses to similar circumstances.
0341 But, can the word, “constellate”, also label the coming together of modules within an archetype as an empedoclement?
Cellulose is composed of glucose molecules that are beta-linked to one another. Starch is composed of glucose molecules that are alpha-linked to one another. Beta-linked polysaccharides cannot be digested. Alpha-linked polysaccharides are easy to digest.
0343 Imagine a bacteria that has a receptor capable of clinging to cellulose, but cannot break the beta-linkage. It binds because exposed cellulose associates to plant damage and plant damage releases various nutrients, including alpha-linked polysaccharides. This bacteria is an opportunist.
0344 At this point, I find it hard to predict what happens next. A biosemiotic reality is going to unfold, but I don’t know how.
I do know that phenomena of this biosemiotic constellation objectify the following noumenal overlay. If I want to scientifically study this specific bacteria, then the real elements of the noumenal overlay give rise to phenomena. Phenomena may be observed and measured. The observations and measurements may be modeled. The models account for the contiguities.
0345 Now, this prokaryotic bacteria is not the only living thing that can take advantage of sites of plant damage. Wood-eating insects do the same. They chew into wood, not because they can digest the beta-linked cellulose, but because they get… well… the same nutrients that the cellulose-binding bacteria want to feed on.
Wood-eating insects, who create the damage that attracts the cellulose-clinging bacteria, inadvertently ingest these bacteria, who do not create the damage that nourishes them. Bacteria are only present to exploit a long-established relation between exposed cellulose and food.
So, when a cellulose-clinging bacteria gets ingested, it can serve as food for the termite (if it dies) or it can simply pretend that nothing significant has changed (if it lives). Bacteria can still hold onto cellulose in the termite’s gut and compete for food with the termite’s own digestive system.
0347 The bacteria’s competition with the termite’s own capacity to digest alpha-linked polysaccharides presents a signaling error. It is as if exposed cellulose (SVs) no longer indicates that food is in the vicinity (SOs) according to the ways that this bacteria interprets the world (SIs).
0348 The specifying sign-relation fails because the bacteria clings to cellulose inside the wood-eating insect’s gut, but this no longer indicates that alpha-linked polysaccharides are available for food.
0349 Then what happens?
Well, phase one ends in this hypothetical scenario when the two independent biomolecular capacities that are innate in the bacteria, the ability to latch onto cellulose and the ability to cleave glucose from alpha-linked polysaccharides, get bound to one another. Now, a beta-linked polysaccharide chain may be held by the latching molecule and be conformationally distorted enough that a terminal glucose can be cleaved by the original cleaving enzyme.
0350 The bacteria lives off some of the glucose that it liberates. But, the potential source of glucose has changed from the starchy alpha-linked polysaccharides that both insect gut and bacteria digest to include previously undigestible beta-linked polysaccharides now available to the bacteria. Indeed, the bacteria release more glucose into the gut than what is available from alpha-linked polysaccharides.
The insect already has pathways for transporting glucose from the gut to the body.
The insect uses the free glucose for its own metabolism. All it needs to do is eat more wood and keep transporting liberated glucose out of its gut, so there is no build-up of soluble glucose in the gut and the bacteria keep that beta-link cleaving pathway operating.
0351 In terms of the S&T noumenal overlay, a beta-linked polysaccharide in the claws and jaws of the “holder” and “cleaver” biomolecules is like an emergent being.
0352 Now, natural selection enters the picture. Bacteria with the innovation prosper. Bacteria with improvements on the innovation prosper even more.
Plus, the wood-eating insect comes under natural selection as well.
0353 Phase two of diversification follows.
0354 At this point, I must remind myself that this scenario is hypothetical.
It is a fiction that is more than a confection (an elaborate frivolous construction), even though it contains a confection (a compounding of two things).
To some, this confection (sugary delicacy) may taste implausible.
But, it offers the curious flavor of an empedoclement.
0355 The sign-elements associated to the S&T noumenal overlay conveys the new message.
For the specifying sign, exposed cellulose in the wood-eating insect’s gut2a (SVs) stands for the emergent being of a conformationally-distorted beta-linked polysaccharide held in the jaws of a cleaving enzyme2b (SOs) in regards to bacterial self-governance3b operating on potential courses of action1b (SIs).
For the exemplar sign, the emergent being of a beta-linked polysaccharide bound to “the claws and jaws” of new complex2b (SVe) stands for a seemingly endless source of glucose from ingested cellulose2c (SOe) in regards to what certain bacteria in the insect’s gut3c are capable of doing1c (SIe).
0356 Each of the sign-vehicles and the sign-objects have observable and measurable facets that will eventually go into scientific models. The scientific models will assist in accounting for each of the sign-interpretants.
Biosemiotic research may be conducted after phase one is complete.
Biosemiotic research can never tell whether phase one is proceeding.
This is the way of empedoclement.
0357 It seems that the wood-chewing insect, so empowered, would go berserk and start to eat every living plant.
Like ripples in still water, biosemiotic waves propagate.
If these wood-chewing insects go after a living plant, then all that plant needs to do is develop a toxin that poisons those little beta-linkage breaking bacteria. Problem solved.
Okay, so these new insects are not invulnerable?
0358 Tell that to the exterminator.
The exterminator facing a wood-eating insect infiltration, knows that certain principles apply. He assesses signs. Is communication (section 10.3) an issue here? I suppose that depends on who is doing the talking and who is receiving the message.
0359 Peirce’s definition of a sign as a triadic relation is crucial for biosemiotics. So, are Peirce’s three categories. Even before getting to a question of communication, there is a character to the S&T overlay that embodies the relational structure of specifying and exemplar sign-relations.
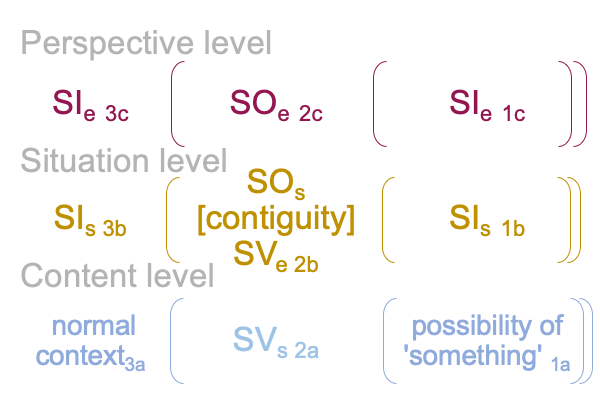
Here is a picture.
0360 These sign-elements, in turn, represent locations in a three-level interscope. An interscope is a category-based nested form composed of category-based nested forms. Each nested form exhibits a normal context3, actuality2 and potential1. Then, the nested forms compose the three levels of contenta, situationb and perspectivec.
See A Primer on the Category-Based Nested Form and A Primer on Sensible and Social Construction, by Razie Mah, available at smashwords and other e-book venues.
0361 I will not show a picture of the three-level interscope.
Instead, here is how the elements of the three-level interscope correspond to elements in the S&T noumenal overlay.
0362 For the specifying sign-relation, a content-level actuality2a (SVs) stands for a situation-level actuality2b (SOs) in regards to a situation-level normal context3b and potential1b (SIs).
For the exemplar sign-relation, the situation-level actuality2b (SVe) stands for a perspective-level actuality2c (SOe) in regards to a perspective-level normal context3c and potential1c (SIe).
0363 In terms of communication, I can imagine that the SVs is like a transmission sent and the SOe is like the transmission received.
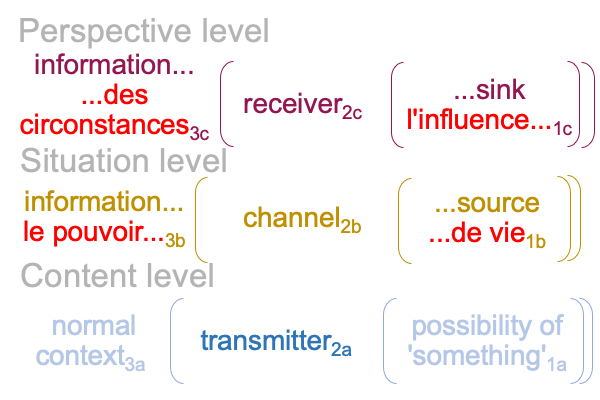
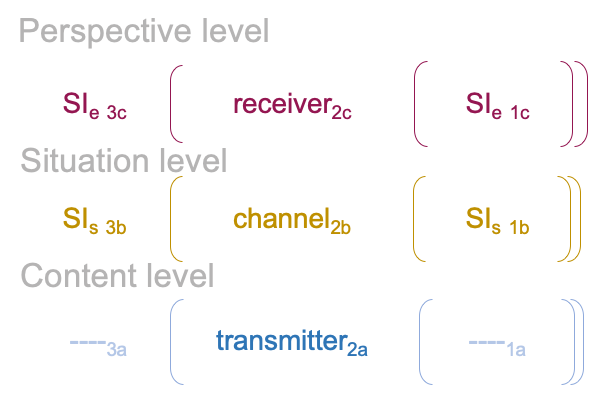
0364 According to Shannon’s information theory, which offers a mathematical model for “information” occurring in human-mediated electro-magnetic transmissions, there are five components to the whole process: (i) an information source, (ii) a transmitter, (iii) a channel, (iv) a receiver and (v) a destination that serves as an information sink.
0365 Can I draw associations to the S&T noumenal overlay?
I suppose so.
0366 The specifying sign-interpretant3b,1b (SIs) is like an information source (i) and the specifying sign-vehicle2a (SVs) associates to the transmission (ii) of something2a that stands for information2b (SOs). Maybe, I can say that the SVs is like a transmitter (ii) and SOs is like that transmission entering the proper channel (iii).
The SOs is contiguous with SVe, so a transmission in the proper channel2b (iii) constitutes an exemplar sign-vehicle2b(SVe) that stands for a perspective-level actuality2c (SOe) (iv), according a perspective-level normal-context3c and potential1c (SIe) (v). So, the SOe may be labeled as a receiver (iv) and the exemplar sign-interpretant3c,1c (SIe) associates to the information sink or destination (v).
0367 Here is a picture.
0368 Of course, my associations are both evocative and fallible.
Here are three implications (F, G, H).
0369 First (F), Shannon’s theory of communication regards biosemiotic sign-interpretants as sources (i) and receivers(iv). These are precisely the elements in the S&T noumenal overlay that need to be explained. Shannon’s theory places them in black boxes and treats them as givens.
What does that imply?
From the point of view of biosemiotics, I wonder, “What is Shannon’s theory actually modeling?”
0370 After all, if I go to the start of the chapter, the two crucial aspects involved in biological change, as characterized by the premoderns, correspond to sign-interpretants.
0371 Shannon’s theory of communication takes these two primordial aspects for granted, in order to concentrate on another question.
What is the channel’s capacity to carry information?
0372 What is so important about the channel2b (iii)?
Hmmm. I see that the channel2b corresponds to “information2b” and to SOs[and]SVe.
What is that about?
Is [and] the same as [contiguity]?
Is this topic about to get fuzzy?
0370 Second (G), Shannon’s theory of communication, as depicted above, is consistent with a biosemiotic approach in regards to the sources of phenomena for communication: transmitter (SVs) (ii), channel (SOs [and] SVe) (iii) and receiver (SOe) (iv).
Okay, so everything’s fine.
0371 Does that mean that Shannon’s theory of communication somehow is relevant to the hypothetical scenario where the... um… vehicle of a beta-linked polysaccharide2a (SVs) is “transmitted” then “received” as the object of glucose molecules ready to power a wood-eating insect’s metabolism (SOe)?
In this case, does the term, “information2b“, correspond to “the ability of a gut bacteria’s dual-modular claw and jaw apparatus to cleave the terminal glucose of a beta-linked polysaccharide”?
It makes me wonder about the meaning, the presence and the message underlying the word, “information2b“.
There seems be a lot going in within the situation-level actuality2b, which is where the SOs transits into the SVe.
0372 Third (H), Shannon’s theory of communication, expressed as an application of the S&T noumenal overlay,presents a simplification that drags the inquirer to that confounding channel2b.
After all, the channel (iii) goes with phenomena.
In human electronic communication technology, the channel2b can carry only so much information2b. Noise is the loss of information2b. Is “noise” information that can no longer be properly received? Or is “noise” some lacking that makes information no longer salient?
Questions like these, as problematic as they are, seem well-cogitated compared to ones that arise when Shannon’s theory of communication is transferred from the dyadic S&T noumenal overlay to a three-level interscope containing the specifying and exemplar sign-relations.
0373 Recall the unexpected and twisted path from point 0270 to point 0300?
Now, maybe I should start getting used to that storyline.
0374 Why?
It’s the story of the evocation of thirdness by firstness.
0375 Initially, Sharov and Tonnessen’s noumenal overlay associates to the specifying sign relation.
Once the Deacon and Tabaczek interscope for emergence enters into the picture, the S&T noumenal overlayincorporates the exemplar sign relation.
0376 Initially, Sharov and Tonnessen’s noumenal overlay belongs to what is of the Positivist’s judgment. It belongs to firstness. Yet, it touches base with secondness, in so far as its own dyadic structure serves as a guide for discerning what should be regarded as phenomena and what is in need of being modeled.
This makes sense, in so far as biosemiotics is the study of the relational thing that all biological entities have in common.
0377 I mean really, how is a biologist going to examine a sample of the relational thing that all biological entities have in common?
Hey, who left a biosemiotic slide in the microscope?
Is it Shannon?
Let me take a look.
0378 Yeah, it’s Shannon’s alright. It has information theory written all over it.
But now, the associations are even more evocative.
0379 The two tendencies that the premoderns identified, the force of life and the influence of circumstances, now associate to normal contexts and potentials. The stuff of strife (situation-level) and love (perspective-level) associate to both thirdness and firstness. These are the things that biosemiotics is supposed to account for.
0380 Shannon’s information theory fixates on the virtual nested form in the realm of actuality.
0381 The perspective-level actuality2c of a receiver2c virtually brings the situation-level actuality2b of a channel2b into relation with the potential of a content-level information transmitter2a.
Yes, these actualities manifest phenomena that can be observed and measured. Shannon’s information theory assists in modeling those observations and measurements. But, do those models tell me about the situation- and perspective-level normal contexts and potentials? Do they tell me about the specifying and exemplar sign-interpretants?
If they do not, then is Shannon’s information theory biosemiotic?