
0659 In section 14.3, the authors report evidence that supports the hypothesis.
0660 What do bacterial researchers observe?
The authors provide examples “hyperstructures” in Table 14.1. These include cytoskeletal filaments, structures made from enzymes, microcomponents, membrane-associated structures, external appendages, DNA-containing structures, phase separation condensates and more.
0661 What do the hyperstructures tell the researcher?
Hyperstructures vary depending on how much energy is available… oh, I meant to say… nutrients are available.
So, the authors propose two temporal phenotypes, corresponding to the appearance of hyperstructures that use lots of energy (NE, non-equilibrium conditions) and ones that conserve current integrity (E, equilibrium conditions).
0662 Here is a picture.
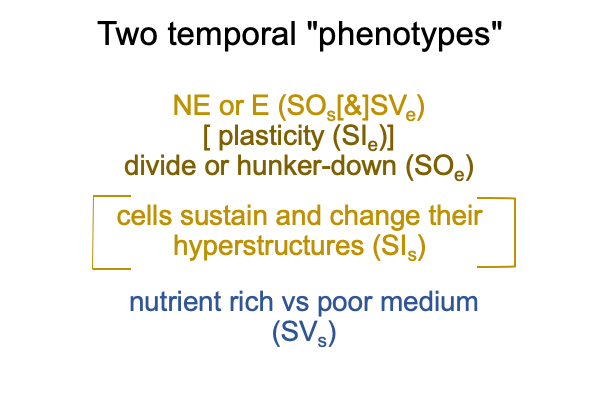
0663 As one might suspect, the two models of hyperstructure maintenance (SIs) and plasticity (SIe) work in tandem. Some transitions from one temporal phenotype to another are easy. Some are not so easy.
As one might have anticipated, a tremendous amount of scientific research has gone into almost all aspects of the hyperstructures, especially in regards to genomic regulation of bacterial DNA. Empirio-schematic research abounds. The problem is that research discoveries appear to have little in common. It’s the problem of inquiry into each leaf occludes the reality of the tree.
0664 The authors provide an example, in section1 4.3.2, for the case when lactose is among the nutrients.
The lac operon, when expressed, contributes to NE hyperstructures. Lactose… transformed into one isomer, allolactose… induces synthesis of the lac operon by binding to a LacI protein that clings to bacterial DNA and represses genomic expression. Allolactose binds to LacI protein and changes it conformation. The allolactose-bound LacI protein lets go of the strands of DNA that it is holding on to, providing the opportunity for formerly suppressed genes to be converted into mRNA.
In sum, for bacteria, lactose is both a nutrient and a cue (SVs) that ultimately says, “Fatten up and divide (SOe).”
0665 Section 14.3.4 describes cell-cycle hyperstructures and signaling. That is to say, “The NE route.” The density of scientific detail stuns the reader. By my estimate, every two sentences summarizes a doctoral thesis. In some paragraphs, every sentence boils down years of research by a doctoral student. The section reads like a biochemical textbook.
After this tour de force of supporting evidence, the authors propose (in section 14.4) that semiotics can be used to explain cues and signals (SVs) in bacteria.
0666 This is where the authors stand today, in 2024, at the threshold of appreciating the um… message, presence and meaning… that biosemioticians observe and measure, in order to construct models of [being alive] and [sentience].
0667 As noted earlier, message (SVs), presence (SOs [&] SVe) and meaning (SOe) go with phenomena.
[Being alive (SIs)] and [sentience (SIe)] are what need to be modeled.
0668 What does this suggest?
The authors do not need to call for semiotics to explain cues and signals in bacteria.
Cues and signals are already specifying sign-vehicles (SVs) in the specifying sign-relation.
0669 Phenomena do not need to be explained.
Phenomena need to be observed and measured by humans who innately recognize sign-elements. Plus, in our current Lebenswelt, we can label those sign-elements through explicit abstraction.
Biosemiotic observations and measurements (what is, firstness) are used to build models (what ought to be, secondness) within the purview of a scientific disciplinary language (relation, thirdness). Such is the empirio-schematic judgment.
0669 Biologist Jakob von Uexkull intuitively senses this. He does not have the advantage of Peircean diagrams of judgment or interscopes. He cannot formulate the category-based nested form, where the normal context of definition3brings the actuality of a spoken word2 into relation with the potential of ‘meaning, presence and message’1. But, he intuitively senses that definition3, spoken words2 and the potential of explicit abstraction1 fit into the conceptualization of semiotic agency. That is why he frames his research in terms of theoretical biology and theories of meaning.
0670 Alexei Sharov and Morten Tonnessen, in their 2021 book, Semiotic Agency, also intuitively grasp this. Unlike von Uexkull, they have loads of scientific research into biological systems, from the micro to the macro, at their disposal. However, like von Uexkull, they do not have the disciplinary language to articulate… um… diagram… their noumenal overlay.
The only way to get to the point of picturing a noumenal overlay is to use diagrams of triadic relations. One value of this examination comes precisely in the act of diagramming the authors’ arguments using triadic relations. Another value of this examination comes in the act of framing the history of biosemiotics as a historical response to the Positivist’s judgment.
0671 Yes, we stand on the shoulders of giants.
Charles Peirce (1839-1914), Edmund Husserl (1859-1939), Jakob von Uexkull (1864-1944), Thomas Sebeok (1920-2001) and John Deely (1942-2017) are giants upon whose shoulders both the authors (and Razie Mah) stand.
Nonetheless, diagrams can really be useful.