0246 Eight years after Mark Twain publishes Innocents Abroad, Henry James (1843-1916) publishes The American.
0247 The year is 1877.
One author represents the attitude of the west, which is still the frontier. The other author expresses the attitudes of the east, which looks across the ocean… or is it the Jordan River?… at the civilization that Americans left behind.
0248 In Chapter 30, Jones discusses Henry James’s novel in detail.
In Chapter 31, Jones turns his attention to Henry Adams (1838-1918), the Old Man of the East, who is not Henry James. The switch of focus is a little disorienting, because both men are named “Henry”.
0249 But, the year is still 1877.
In this year, federal troops withdraw from the South, ending Reconstruction. The new post-war era is to be called, “Redemption”. Three decades later, the war itself would get a new label, “the Civil War”. It is amazing what a change in labels can accomplish.
In this year of 1877, the American labor movement surfaces with a surprising railroad strike.
In this year, a descendent of President John Quincy Adams, Henry Adams, resigns from his professorship at Harvard, disappointed that his dream of a unique American Mandarin class had not come to fruition. If he had only lived another half of a century. Once science encourages the birth of “expertise”, Henry Adam’s dream becomes a reality.
He must have known what was coming, because he writes an autobiography that is published right after his death and after the conclusion of the so-called “First World War”.
Jones tells the story of unfortunate Henry.
0250 The War of Southern Rebellion and the War of Northern Aggression derails the American experiment, as well as Henry Adam’s career as a public intellectual. Even his professorship at Harvard does not seem to give him a lift.
0251 Chapter 33 covers what happens to Henry Adam’s wife, and how Henry spends his end of days.
At this juncture, I would like to mention an American philosopher whose life (more or less) overlaps with Henry Adams. This philosopher is Charles Sanders Peirce (1839-1914). Charles is the son of a Professor of Mathematics at Harvard University. Tremendous intellectual ferment occurs during the latter half of the nineteenth century. Very little of that turmoil influences the westward expansion of the Re-united United States or the migration of Catholic Europeans to the East Coast (Italians join the incoming Irish and Germans).
Peirce does not concern himself with Protestant or Catholic concerns. Rather, like Henry Adams, he recognizes that everyone is talking as if science is the key to all knowledge worth knowing. The century-long contests between Catholic and Protestant doctrines are no longer relevant, since science surpasses both. Unlike Henry Adams, he reads the Baroque scholastic, Francisco Suarez (1548-1617), then arrives at the same definition of sign-relation as John Poinsot (1589-1644).
So, while Henry Adams ends his days writing paeans (in effect, aesthetic travel guides) to Catholic European tourist destinations, Peirce wrestles with the causalities inherent in sign-relations. The diagrams in this examination are products of Peirce’s postmodern reclamation of Baroque scholastic thought.
0252 The student should read chapters 35, 37, 38, 39, 40 and 41.
0253 Chapter 35 discusses Theodore Roosevelt (1858-1919), who serves as president at the turn of the century (1901-1909). His presidency starts with the assassination of William McKinley. At the time, Roosevelt is Vice President.
Bullets are in the air during this time. Lakota leader Sitting Bull (dies 1890), New Orleans Police Chief David Hennessy (1890), Lamatta Chief White Bird (1892), US Chicago Representative Carter Harrison Sr (1893), President of France Marie Francois Sadi Carnot (1894), King Umberto in Italy (1900), and so on.
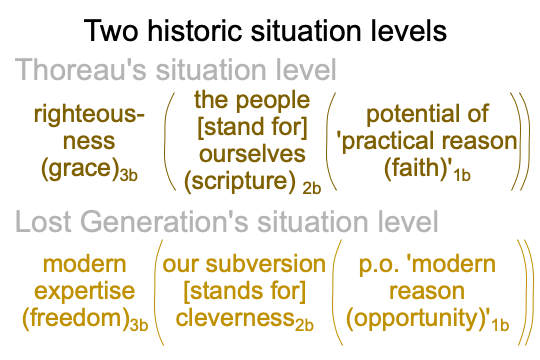
0254 Thoreau’s option reaches a nice optimum, with the people2bm remaining Protestant as they take the form of democracy2bf. The people2bm amend the Constitution, now that the states are reunited and… um… redeemed. Americans are still walking3a to settle the frontier, but now they can take trains out west. Some Americans are driving automobiles. Some are taking mechanized passenger ships back to Europe, for “vacation”.
So, walking3a stands in for what Thoreau does, that is, ideating reality3a.
Here is a picture of Thoreau’s option, as the western frontier comes to a close.
Notice that ‘pure reason (innocence)’1a occupies the same slot as ‘gun’1a. By not carrying a gun1a, one signals one’s virtue2am (and the nature2am that unites us). By deploring people who carry guns, one denounces their lack of virtue2am.
What does this imply?
When the people2bm [stand for] themselves2bf, the selves2af that the people2bm stand for are the virtuous ones2af, not the ones who carry guns.
If that makes sense, then you might be a candidate to become an expert.
0255 Teddy Roosevelt promotes sports and sportsmanship. Democracy is so successful that Protestant people2bm will be supplanted by our nation2bm. Plus, our nation2bm stands for “a muscular Christianity2bf“. Muscular Christianity2bf virtually situates our attitudes2af in the normal context of winning3a arising from the potential of ‘fair play’1a.
Here is a diagram.
0256 Notice how ‘fair play (our lack of guilt)’1a occupies the same slot for ‘gun’1a in the original American identity. Teddy Roosevelt is known for his “gunboat diplomacy”. So, guns have not really left the realm of possibility1a. Rather, ‘the gun as a talisman for some opaque transcendental that is too terrifying consider’ retains its charisma, since guns are employed for assassinations by criminals (1890, maybe 1893), anarchists (1894, 1900, 1901, 1914) and agents of the state (1890, 1892).
So, what is the point?
‘Guns’1a have no place in ‘fair play’1a. At the same time, ‘guns’1a serve as indicators for a lack of fair play1a, so to speak. The use of ‘guns’1a defies practical reason1b in a nation2bm that stands for Christianity2bf in the normal context of expertise3b.
0257 Chapter 37 discusses an example of how expertise3b develops at the turn of the century. The artisan is eclipsed by the industrial designer.
0258 Chapter 38 discusses the closing of the American frontier in the popular (or… rather, the literary class’s) imagination. A key event is the Columbia Exposition in Chicago in 1893. Jones reports on a speech by Frederick Jackson Turner at this spectacular “World Fair”. He announces that frontier opportunities had come to a close.
According to Jones, Theodore Roosevelt personifies a new American identity, characterized by the above diagram. Situation-level matter2bm veils the federal government, as a centralizing state, beneath the tapestry of nationalism2bm. Our nationalism2bm stands for a strenuous Christianity2bf in the normal context of expertise3b and arising from the potential of practical (that is, “not religious” reason)1a. Protestant Christians2bf virtually situate the sportsmanship2am that trains healthy attitudes2af.
0259 At the same time, Protestant Christians2bf increasingly become wage slaves to a managerial class3b(1b) working for oligarchs2c, such as the Mellons, the Rockefellers and the corporations that they own.
0260 Part 2 (The Rehabilitation of “Nature”) leads to Part 3 (The Return of the Satanic Wilderness).
The first wilderness stretches out from the fortress edge of the Massachusetts Bay Colony.
The second wilderness stretches to the north (Canada), the south (Mexico and Central and South America), the east (across the Atlantic to Europe and the Middle East) and west (across the Pacific to the Far East).
That sounds like an empire to me.
The second “wilderness” is tamed by the American empire.
0261 Chapter 39 discusses the writer, Jack London (1876-1916). London comes to age in San Francisco. He reads European authors of the day. He circulates in Bohemian circles. His novel, Sea Wolf (1904), serves as a witness to Darwinism replacing Calvinism as the hidden grammar of the nascent American empire.
I wonder, “Where have I seen the term, ‘hidden grammar’, before?’
Oh, it2a labeled the contiguity in the content-level actuality2a.
I wonder, “Can I adjust Teddy Roosevelt’s revision to embrace Darwinism?”
This serves as a good template for reflecting on the author’s review of Sea Wolf.
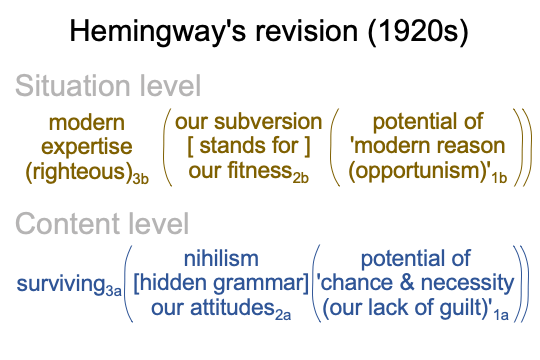
0262 Chapter 40 discusses the writer Ernest Hemmingway (1899-1961), who was born in Oak Park, Illinois. That is a nice suburb of Chicago.
Henry Adams dies in 1918, when the First World War ends. Adams’s autobiography, The Education of Henry Adams comes on market. At the same time, Hemmingway is recovering from wounds that he received while driving an ambulance in the First World War. After the war, he makes his way to Paris, where he meets Ezra Pound and other literary figures of the so-called “Lost Generation”.
0263 Jones sets the scene by discussing the birth of modern art. “Modern art” involves a clever and subversive re-definition of the term “visual arts”, from decorative to… well… transgressive.
I suppose this indicates that neither Protestantism, nor nationalism nor Darwinism are adequate to appreciate the end of the world as everyone knew it.
Oh, I meant to say “the First World War”.
0264 What is left?
Science, of course. Modern expertise3b carries a certain righteousness3b, as well as certain freedoms3b. Modern reason1b accesses opportunities1b that practical reason1b does not. For example, consider the logics of modern finance, quantum mechanics, industrial-production, and mass-marketing. All these fields develop in the interwar period, along with the ideologies of Marxism and Fascism.
0265 What do these ideologies have in common?
They both rely on modern experts3b operating of opportunities1b.
Of course, their subversions2bm are um… “value neutral”1bf.
0266 Here is how I associate Hemingway’s “Lost Generation” approach to Jack London’s revision.
0267 Chapter 41 covers William Falkner (1897-1962), who writes a novella titled, “The Bear”.
This portion of Jones’s book looks back at the American identity and wonders, “How have we sold our identity?”
0268 It seems that Americans sold their identity3b for a bowl of porridge.
Or something like that.
Americans pass from sola fide3b, sola sciptura2b and sola gratia1b to the following nested forms, as well as everything in between.
0269 The student should read Chapters 43, 44, 46 and 47.
0270 Chapter 43 opens with a quick synopsis. By 1902, the American frontier had been replaced by eye-catching progress. Also, popular novels fictionalize the failure of Protestant America to envision a common identity.
This makes sense, in so far as, in the Protestant movement, every literate person is granted equal disposition1a to read the Bible2am, and render his own interpretation2af. That interpretation2af generally falls within established congregations (the Puritans, the Methodists, the Presbyterians, and so on). The American Revolution accommodates the various powers1b by establishing a relatively weak tripartite central (or “federal”) government over strong state governments1b. This political arrangement is a practical solution1b, allowing people of certain faiths1b the freedom3b to self-segregate into their own jurisdiction2b.
0271 Here is a picture of (what I will call) the Virginian’s option.
0273 What is this potential of ‘gun’1a?
It is easy to understand how the Bible (for Christians) or nature (for Transcendentalists)2bm unites individuals of like minds2af.
But how does the ‘gun’1a potentiate that actuality2a?
0274 Well, when one is walking3a with a ‘gun’1a, one is conveying a meaning1a, a presence1b and a message1c.
The meaning1a has something to do with the possibility of self-defense1a, especially in regards to mortal conflict. A gun1a cannot help finding water. A gun1a can help when a rustler is “guarding” the water hole.
The presence1b has something to do with the possibility of practical reason1b. Practical reason1b allows the people2bm to stand for themselves2bf, in the normal context of freedom3b. Practical reason1b is another label for ‘responsibility’1b. Recall that freedom and responsibility constitute a “co-opposition”. When one is the normal context3 the other informs the potential1. Responsibility associates to faith. Freedom associates to grace.
0275 The message1c?
That is a good question, since the perspective level does not even appear in the above figure.
I can say the following from the clues offered in the examination.
0276 Whatever the perspective level is, I must assume its normal context.
In other words, a perspective-level “assume3c” virtually brings a situation-level identity3b into relation with the potential of content-level structure3a. The logics of thirdness are exclusion, complementarity and alignment.
In the Virginian’s option pictured above, I must assume3c that situation-level freedom3b (or grace3b) situates content-level walking3a (or ideating reality3a).
0277 More crucially, I assume3c the perspective-level actuality2c.
Of course, the most magnificent assumption3c pertains to the object that brings all into relation2cC. That2cC includes “God”. And, perhaps I can add, “us”, as participating in this object2cC.
According to the twentieth-century German philosopher and historian, Eric Voegelin (1901-1985), Christians come up with the most differentiated expression of the society-tier perspective-level actuality2cC, sometime between the 600s and 1000s, centuries before the so-called “Reformation”.
0278 Finally, I somehow intuitively grasp the potential1cC that there can be an object that brings all into relation2cC.
I wonder whether this is where the gun1a comes into the picture, as a talisman for consilience1c in the face of a failure of practical reason1b. When practical reason1b fails, then the people cannot stand for themselves2b in the illumination of their freedom to assemble3b, and live3b, and express themselves3b.
0279 Where are we to go when people who live without a sense of guilt1aactualize attitudes2af that express the hidden grammar of nihilism2am in the normal context of their survival3a?
It is a jungle out there.
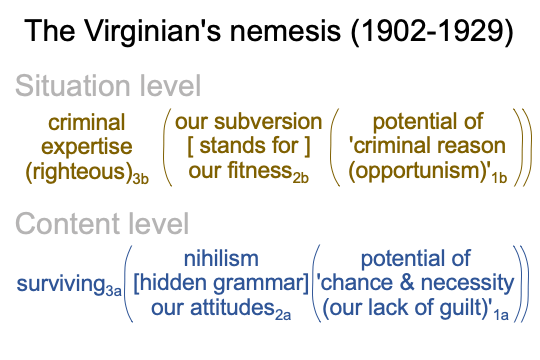
0280 Here is a diagram containing that question.
0281 On the situation level, the normal context of criminal expertise (the righteousness of criminals and their lawyers)3b brings the actuality2b, {the principles of our subversion2bm [substantiate] our fitness over our victims2bf}, into the possibilities inherent in ‘criminal reason, which is opportunistic to the core’1b.
0282 In Chapter 44, Jones looks at The Virginian, a novel by Oren Wister (1860-1938).
The book is published in 1902. The movie comes out in 1929.
0283 In Chapters 46 and 49, Jones considers movies starring Clint Eastwood (1930-present).
0284 Chapter 42 reports on Walter Lippmann (1889-1974), born in a wealthy Jewish family in New York. He receives a classical education at a private school. In 1922, after America’s victory in the First World War, “the war to end all wars”, he publishes a book titled Public Opinion.
Perhaps, he knows that the similarly positioned Edward Bernays (1891-1995), nephew of renown psychotherapist, Sigmund Freud, is hot on the same topic, publishing Crystallizing Public Opinion (1923) and Propaganda (1928) in quick succession. After the Second World War, Bernay capitalizes on his fame by writing Public Relations (1952) and The Engineering of Consent (1955).
0285 I like to call the First World War, 1914-1918, the First Battle Among the Enlightenment Gods, The War Among Naive Mercantilists.
During this time, Lippman works for the intelligence section of the Allied Expeditionary Force in France. He helps draft President Woodrow Wilson’s Fourteen Point Speech. He hears about the success of George Creel’s Committee on Public Information, America’s first propaganda ministry. Creel (1876-1953) later wrote a tell-all titled, How We Advertised America (1920).
0286 Lippmann concludes that identity3b may be framed as a typology, based on (what E. Michael Jones calls) categories of the mind.
Plus, the subtlest and most pervasive influencers create and maintain these categories, using broadcast media.
0287 Now, I pause to remind myself of the contemporary Thomist construct of identity3b and structure3a in order to consider the situation and content levels actualities that get packaged into a stereo- (two) -type (actualities).
Here is a diagram.
What is a stereotype?
The first type in “stereotype” dwells in content-level matter2am. It2am substantiates a form2af, corresponding to the way that we experience the world2af. We are structured3a to be disposed1a to certain attitutes2af that are informed by one or another stereotype2am. The task of propaganda1b is to situate our attitudes2a by placing a typology in content-level matter2am.
Then, the form2af that we have been structured3a to experience2af is situated according to a principle of unity2bm that substantiates an embodiment2bf. The task of propaganda1b is to prime2bm a response2bf through the second type of “stereotype”, dwelling in the situation-level principle of unity2bm.
The two types in stereotype play the same music through different level matters.
The result sounds so real.
0288 Lippmann writes at the dawn of popular acceptance of moving pictures.
0289 Moving pictures?
In the realm of image processing, a sequence of slightly different images2am substantiate the form of a mechanically driven display of one image following the other2af. Then, the human mind has the power to apply a principle of unity2bm that substantiates the embodiment of motion2bf.
Then, sequences of moving-picture clips2am may be strung together2af and the audience will apply another principle of unity2bm to appreciate a scene2bf.
This constitutes the first type in “stereotype”.
Scenes portray the content-level matters2am of action and plotline2af, and the audience spontaneous situates the content-level form2af using an implicit principle of unity2bm into a cognitively embodied “narrative”2bf.
This constitutes the second type in “stereotype”.
The result looks so real.
0290 It is almost as if identity3b, powers1b, structure3a and dispositions1a are simply going along for the ride.
Why would this be the case?
Is this how humans evolved to be?
0291 So, what happens when stereotypes are promulgated (through cinematic narratives manufactured in order to serve… how shall I say it?… “special interests”)?
Indeed, Jones wonders whether Ernest Hemingway and the Lost Generation are stereotypes manufactured by Madison Avenue (New York) and Hollywood (Los Angeles) as expressions of American cultural imperialism.
A new system arises in order to manipulate public opinion2bm and orchestrate the accumulation of power and money2bf through the invention and the promulgation of stereotypes1am.
The vessel3b(1b) that accrues both power and money is a network of big(il)lib power brokers who fashion themselves as “liberal” (that is, freedom-oriented) but are really interested in using the authority of government to suppress anyone who would hold them to moral account (that is, responsible). In other words, this vessel is a contradiction in terms, being simultaneously liberal and illiberal.
Consequently, I label this network, “big-government (il) liberalism”, or “big(il)lib” for short.
0292 Here is a picture of Walter Lippmann as a nemesis to the American tradition of walking with a Bible and a gun.
0293 On the situation level, the normal context of working the system3b brings the actuality2b where {our success in accumulating money and power2bm stands for our fitness over the goyim2bf} into relation with the potential of using propaganda1b as a style of public reasoning1b that cultivates the hidden grammar of stereotype2am-fueled attitudes2af.
On the content level, the normal context of manufacturing consent3a brings the actuality of propaganda1b-situated stereotypes2am substantiating public attitudes2af into relation with the potential of ‘selling an agenda, as if we do not benefit from moving the product’1a.
Jones frames this nemesis as an inversion of Jeffersonian democracy, where the people stand for themselves2b.
0294 Chapter 45 mentions Ayn Rand (1905-1982), the author of Atlas Shrugged, (1957), as a teaser for his real intent, which is to discuss the fates of the movie-star, Gary Cooper (who plays the role of The Virginian in the movie) and the literary-star, Ernest Hemingway (who acts as an author of American literature as the tradition turns into a curation by the mavens of modern art).
Does it transform into some disorienting house of mirrors.
Does propaganda constitute the new American written word, just like the moving pictures?
0297 Here is a diagram of how this new literary style might operate.
A new American literature2bf serves as the embodiment2bf of big government (il)liberalism2bm.
0298 Chapter 47 discusses The Moynihan Report.
In 1965, the Catholic Assistant Secretary of Labor, Daniel Patrick Moynihan (1927-2003), releases a report concerning why blacks (a category of the big(il)libs) are not as economically successful as whites (another category of the big(il)libs). The primary reason is broken families (which are the products of a separate big(il)lib agenda, called “sexual liberation”). Sexual liberation produces broken, fatherless, families, which then must be supported by federal programs with titles such as “welfare for families with dependent children” (a big(il)lib program). The result is a cycle where the children of welfare families end up as… um… welfare-seeking heads of families with dependent children.
0299 Jones has already written extensively on this topic.
But, does he realize that the public responses to this report constitute a new genre of American fiction?
0300 The Moynihan Report is immediately opposed by big(il)lib establishment figures such as Robert W. Spike (1923-1966), then executive director of the National Churches’ Commission on Religion and Race. Author of The Freedom Revolution and the Churches, he calls for a political solution. The “whites” need to expiate their guilt. Or, something like that.
0301 To me, if this doesn’t sound like a new American fiction, I don’t know what does.
Pity the reverend who has a control file at the disposal of someone with money and power.
Pity the fellow who speaks up, then is served a lawsuit about some… how shall I say it?.. episode of sexual liberation, whether true or not.
0302 This is the literary genreof manufacturing consent.
Stereotypes2am are like esse_ce. They constitute a new (virtual) wilderness where human dispositions1a are subject to literary and cinematic manipulation. Our attitudes2af are like the essence that stands between human structure3a and our evolved dispositions1a.
The content-level category-based nested form with the normal context of making influence3a is also like esse_ce. Making influence3a is the normal context for a cognitive wilderness2a arising from the potential of ‘selling a product or agenda’1a. Both the normal context3a and the potential1a are sign-interpretants of a sign-vehicle, a perspective-level actuality2c that stands behind the curtain2b (so to speak) of stereotypes2am, attitudes2af, expert self-regard2bm and the literature of… what?
What is the appropriate label for this new American genre?
0303 Chapter 48 concerns critical race theory and the destruction of American identity.
Can I locate the sought-after label within these concerns?
0304 Jones writes about the subversion of Catholic identity in the ethnic neighborhoods of Philadelphia.
The intelligensia2bm – the literary classes – now refer to the people who used to live in those neighborhoods as “white”2bf. The unspoken accusation is that these “whites2am” are the reason why “blacks2am” cannot make a living. It is their attitudes2af. The unspoken accusation covers up the reality that the federal government’s welfare programs have subsidized broken families for generations.
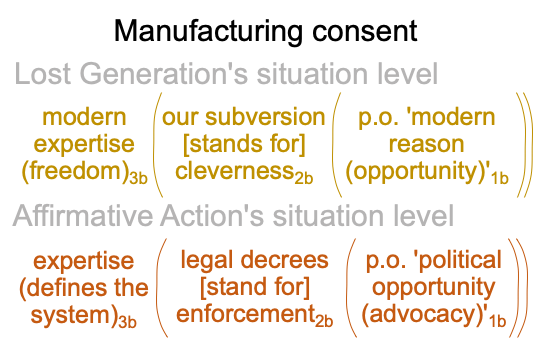
0305 The only promoted remedy is for the federal government to step in and take affirmative action. This generates new expertise and constitutes the (il) of big(il)lib. Freedom of thought for modern expertise3b is mirrored by an expertise that defines the system3b through the fiction of legal decrees standing for enforcing morals2b according to the dictates of ‘an opportunistic modern situational ethics’1b.
0306 Here is a comparison of the Lost Generation’s and Affirmative Action’s situation level nested forms.
Each term seems to be a variant of the other.
What happens when both these situation-levels are reflections in the mirror of… what?…
0307 If Jones states that critical theory is the nemesis of American Identity, then is something else standing in the mirror of this nemesis?
0308 The stereotypes of race are “black” and “white”.
The stereotypes of sex are “victim” and “aggressor”.
The stereotypes of gender are “legion” and “straight”.
0309 One stereotype cannot be labeled.
Why?
It is not a stereotype at all. It is as real as the people with money and power3c, who can make influence3a and impact3a in order to sell a product1a and an agenda1a. These are the hidden ones, who hire the psychologists and sociologists, to survey the people… or rather, their internet usage… in order to know how their interventions2c shape attitudes2af and appear relevant2af.
Surely, they3c are looking for opportunities1c.
0310 These hidden ones are as real as Reverend Dimmesdale and Hester Prynne in Nathaniel Hawthorne’s The Scarlet Letter. They matter.
0311 They finance the types that come in “stereo”, as if there are two speakers, but one musical composition in the air.
Shall I play a game with metaphors?
Two speakers (Q and R) mirror one another.
The first speaker3b (Q) constructs critical theory2bf and occupies a position of academic renown, complete with a title such as Distinguished So-and-So Chair of the Institute for the Investigation of Whatever Stereotype Needs to Be Promoted. This expertise3b virtually situates making influence3a.
The second speaker3b (R) sets the warp and woof of critical theory. Warp and woof? Let’s not get into the looming machinations in which the actions of academic employment are affirmed. These are the speakers3b who, by their very presence in academic institutions, constitute a system of surveillance as formidable as any… well… “clerical” apparatus. This expertise3b makes an impact3a.
0312 Here is the diagram for the identity3b and structure3a of critical theory as nemesis of what it means to be an American (Q). This is the sensible construction of the first speaker3b.
0313 To me, this big noumenon corresponds to experts3b that propose valuations2b that engender opportunities1c for the Relativist One3c, as discussed in a separate three-part e-book by Razie Mah, titled Original Sin and the Post-Truth Condition,
This examiner suggests that the literature2bf of critical theory is a novel genre of American fiction.
Jones’s application of existence (the content level) and essence (the situation level) tells me that making influence3a is like walking3a, the actuality of {stereotypes [hidden grammar] our attitudes}2a is like {the Bible [unifies] us}2a, and the potential of ‘guiltlessly selling product’1a is like the potential of ‘whatever carrying a gun intends to defend against’1a.
0314 Imagine carrying a metaphysical gun1a that would shoot Cupid before he could wound you with one of his self-annihilating arrows of desire1a.
This would be the same metaphysical gun1a that may be used against a critical theorist2b.
0315 But, what about the people who are peirced1b by the arrows of critical theories2bf and then fall in love with the promulgated stereotypes2am, whose hidden grammar makes the critical fictional forms so relevant2af (R)?
It is as if they are trapped in the mirror of nemesis.
0316 Here is a picture of the sensible construction of the second speaker3b (R).
0317 It is as if the believers3b are like guns1a.
They3b are tools that the ones who work the system3b use to promulgate their self-promoting2bm literature2bf.
Believers3b spout the literature2bf of the ones with self-regard2am as if they are legal decrees2bm, demanding sovereign enforcement2bf. These legal decrees2bm pretend that stereotypical fictional matter2am is relevant2af enough to make an impact3a arising from the potential of ‘their adopted agenda’1a.
0318 Believers vicariously share the working experts’ lack of guilt1a.
They don’t need to carry guns1a.
They carry a political agenda1a.
That agenda1a makes an impact3a because operators3c in the hidden perspective level are flush with money and power2c and looking for opportunities1c.
0319 The two diagrams portrayed above constitute America’s contemporary wilderness.
0320 The student should read chapters 50, 51, 52 and 53.
Welcome to a simulated reality constituted by agents that work the system (Q) and minions that believe in the system (R).
The system is always changing, yet always the same, because it lives by cancellation of their opposition. The system replaces a suspect person’s identity3b with a stereotype2am that opposes the adopted agenda1a. The stereotype-tagged guilty one2af deserves social cancellation1a (the gun1a of critical theory).
0322 Jones tells the story of a person who… well… had an old-style American identity.
In the big noumenon, identity3b virtually situates structure3a.
In the big noumenon of the nemesis of the American Identity (Q), big(il)lib expertise works the system3b and virtually situates manufacturing influence3a.
In the big noumenon of those self-identifying within the mirror of nemesis (R), big(il)lib expertise believes in the system3b and virtually situates making an impact3a.
0323 For example, self-regarding racial theorists2bm (Q) compose literature2bf that isolates stereotypes2af that – through an unspoken contiguity called “hidden grammar” – substantiate attitudes2af within the normal context of making influence3a and the potential of ‘selling a narrative (that veils the guilt of interventionalists on the perspective level)’1a.
0324 Oh, did I say “narrative”?
I meant to say, “product”.
0325 For example, educational committees2bm (R) distill what needs to be enforced2bf from the fictions of critical theory2bf. These experts3b advocate policies2b in light of what they perceive are political opportunities1b. In this way, the propaganda1b underlying the literature2bf of higher-level authorities2bm manifests as stereotypes2af that are regarded as relevant2bf in the normal context of making an impact3a operating on the potential of ‘the adopted agenda’1a.
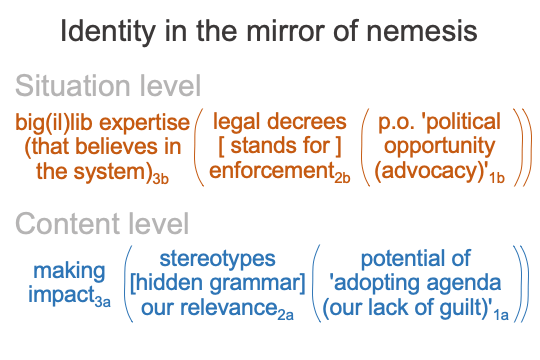
0326 Allow me to compare critical theory (Q) and the mirroring identity of the big(il)lib believer (R).
Here is a comparison that starts with the situation-level normal context.
0327 On the side of nemesis (Q), the normal context of big(il)lib expertise3b has the intellectual freedom to “work” the system. Unrestrained intellectual liberty3b subverts the (old) targeted system (say, the institutional structures3a of the American Identity of the 1900s), using apparently clever tactics (typical of the manipulation of words in propaganda1b).
On the side of the mirroring identity (R), the normal context of big(il)lib expertise3b internalizes the knowledge created by cutting-edge critical theorists3b. Even though both nemesis and mirror are regarded as experts, there is a matter-of-fact distinction. For example, when a language professor teaches a course informed by the critical theories of Foucault3b, she contextualizes the French theorist’s proclamations as if they are legal decrees2bm, carrying the weight of authority2bm. The students know that the professor will use the opportunities afforded by classwork1b to enforce2bf that authority2bm.
0328 The comparison continues in the following figure.
0329 Here is a reminder. “Big(il)lib” is short for big government (il)liberalism. The bigger the government gets, the more illiberal the self-declared “liberal” will act.
0330 Potential on the situation level1b situates the entire content-level nested form, particularly the actuality2a.
0331 On the nemesis side (Q), the technical language of big(il)lib reason1b yields propaganda1b which situates the stereotypes2am that guide our attitudes2af by way of a hidden grammar. [Hidden grammar] is the contiguity between the matter of stereotypes2am and the form of our attitudes2af.
On the side of the mirroring identity (R), the action-oriented language of political opportunity1b yields advocacy (calls to action)1b that situate stereotypes as matter2am and proclaims the importance of the attitudes that stereotypes call for2af.
0332 The comparison is completed in the following figure.
0333 On the nemesis side (Q), making influence3a is situated by the potential of propaganda1b.
On the mirror side (R), making an impact3a is situated by the potential of political opportunity1b.
Anyone tagged with one of the old American identities is outside this system. Indeed, the two sides function to sap the resources of those outside their fiction.
0334 On the nemesis side (Q), liberated theorists3b write the theoretical thought-pieces2bf that govern the stereotypes2am that “speak to” our attitudes2af.
On the mirror side (R), educated believers3b derive the laws inherent in those thought pieces2bf in order to enforce2bf the realness (not fictionality) of the stereotypes2am, thereby making them palpably real2af or “relevant2af“.
Is that not what good fiction is supposed to do?
0335 Finally, both the nemesis of American Identity and those who mirror the nemesis manifest the potential of having no guilt1a, just like the Satan, at the opening of John Milton’s Paradise Lost. And, for Americans, that potential is precisely what carrying a gun aims to mitigate.
0337 The colonies expanded into the wilderness of the frontier west.
The states expanded into the wilderness of nature.
The United States of America expanded into the wilderness of empire.
Now, we, the people, expand into the wilderness of a house of mirrors.
0338 The final three chapters and the epilogue convey a sense of resignation. Is the fiction that critical theorists write to be regarded as a comedy or a tragedy? Or is it a liberation or a trap? Nothing is as it appears to be. Not even the author remains undistorted.
0339 Even though Jones does not label the house of mirrors that constitutes Critical Theory “the new postmodern American fiction”. Jones’s treatment of Joe Ritchey tells of someone who was not only canceled, but ruined, because of his American Identity.
0340 Without explicitly asking the question, Jones poses the quandary, “Do the institutions that support critical theory as a way to turn their world into a fiction have historical precedent?”
Which of the following pairs of comparisons does it parallel?
0341 The student is left to formulate an answer to the question.
0342 My own epilogue begins with a cheer for E. Michael Jones for this thought-provoking exercise.
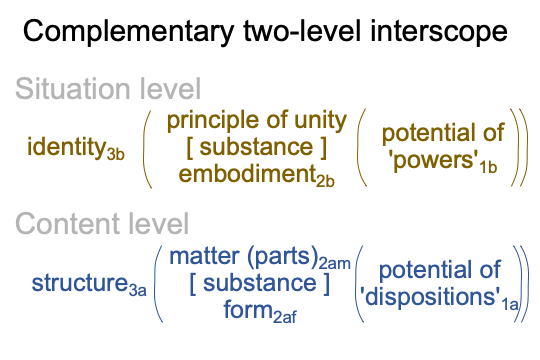
0343 My examination begins with the introduction of a model through which I (the reader in 2026) can appreciate contemporary meaning, presence and message within historic American literature. This is discussed in points 0027-0042.
As it turns out, a big picture of his original model of “existence precedes essence” may be diagrammed as a two-level interscope where identity3b virtually situates structure3a.
0344 Here is a picture.
This examination has been informed by a postmodern version of Jones’s Thomistic insight.
0345 What about Jones’s provocative writing style?
Readers from Catholic backgrounds might have a little difficulty accepting the text as written, because it (sort of) sounds like apologetics. My own difficulty? The model application, although consistent, does not shine, as it does in this examination. Jones applies the model. He does not articulate it. In this, my examination adds value to Jones’s text.
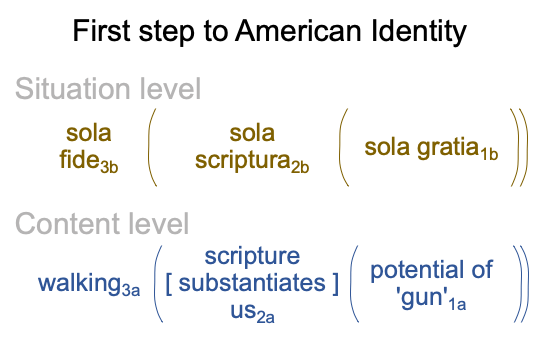
Readers from Protestant backgrounds may have serious difficulties with Jones’s text, in so far as the author does not respect Martin Luther, John Calvin, King Henry VIII and other founders. Plus, there is continuous mention of Satan as the hero of John Milton’s Paradise Lost. Plus, Jones seems to have no fondness for the situational import of sola fide, sola scriptura and sola gratia. This slogan situates the title of Jones’s book.
0346 Indeed, the first historical step to the American Identity contains the Protestant slogan as situation level and the title of Jones’s book as content level, as shown below.
0347 A meditation on this nested form tells me that Jones presents a gift to the Protestant and the American. Here is the first expression of the American Identity conforming to the scholastic ideation that existence precedes essence. Existence associates to the content level. Essence associates to the situation level.
At the same time, the esse_ce of scripture2a is contiguous with the essence of us2a, as Americans to be.
0348 And the gun?
Is it really a talisman for self-protection from those who pretend to be absolved of the doctrine of original sin, that is, from those who deny the fall of man?
Surely, this question will inspire future inquiry.
0349 Overall, Puritanism (Calvinism) ran its course for a century before the American Revolution. Then the romantic rehabilitation of nature ran its course for a century before the Civil War. Then, the nemesis and its mirror of abolitionist and northern aggressor, democratic theorist and voter-rights advocate, secular scholar and value-free administrator, aggrieved theorist and agenda-pushing educator, and (lest I forget) rabbi and jew, has run its course for a over a century.
What is a good title for the coming end of the current saeculum?
Here is my suggestion.
A Shattering of the House of Mirrors.
Or maybe, A Reconstitution of the American Identity, will do.
0350 My thanks to E. Michael Jones for this work. I hope that this examination adds value to his exposition and serves as a companion for the student interested in the topic.
SaH0047 The following essay, originally written in 2016, comments on a book by a Lutheran Pastor about heretical influences in contemporary American society. The author is Peter Burfeind. The title of the work is Gnostic America: A Reading of Contemporary American Culture according to Christianity’s Oldest Heresy (Pax Domini Press, 2014). Please have this book at hand for the full story.
These comments are not a close reading. They situate Burfeind’s text with category-based models. These models were proposed and developed in How to Define the Word “Religion”, plus Primers 1 through 10. They were refined in the Course on Implicit and Explicit Abstraction. They portray relational and logical structures.
Also, my discussion presumes the first singularity. Gnosticism was cultivated in an early ancient civilization. As such, I expect its origin myth to be a fairy tale about the first singularity. The first singularity is discussed in the Crystallizations of the Fall series plus An Archaeology of the Fall, available at www.smashwords.com and other e-book venues.
In short, these comments apply a suite of postmodern scholastic hypotheses to Burfeind’s excellent text. They are different from, yet complementary to, Burfeind’s passionate argument.
SaH0048 This exposition constitutes a strand in the Razie Mah’s online course: Semiotics and History.
The various strands are listed in Razie Mah’s blog for February 3, 2026
‘Words that belong together’ are denoted by single quotes or italics.
0001 Peter Burfeind wrote Gnostic America: A Reading of Contemporary American Culture according to Christianity’s Oldest Heresy (Pax Domini Press, 2014).
This book is composed of four parts. The first part covers the fundamentals of Gnosticism as a Christian heresy. The second and third parts explore cultural manifestations. The fourth part applies these fundamentals and explorations to contemporary American Neo-Evangelical Christian churches.
0002 Burfeind argues that two Gnostic movements contend for sovereign power, progressives and Neo-Evangelical Christians.
To me, big government (il)liberals (BG(il)Ls) beat these Christians to the punch. The federal government has established a religion. It is not Christian. BG(il)Ls run Gnostic bureaucracies where authorities deny that they are religious at all. BG(il)Ls are the self-anointed angels (or more accurately, daemons) of a New Age dawning in America.
Christians who self-describe themselves as “spiritual, but not religious” accommodate BGL’s increasing sovereign power and aggression.
0003 Burfeind is not alone in this awareness. Over 60 years ago, Eric Voegelin expressed similar ideas in his short but significant book, The New Science of Politics.
This raises a question. What is the meaning, the presence and the message underlying the word “Gnosticism”?
With this question in mind, I turn my attention to Burfeind’s text.
Students should read chapter 1 of Burfeind’s text before proceeding.
0004 From Burfeind’s introduction, I know that Gnosticism is a very old heresy.
How old?
From chapter 1, I suspect that Gnosticism predated Christianity. Indeed, the death and the resurrection of Jesusrebuked Gnostic intuition. Why would Jesus retain the evil material shell of His body?
Jesus highlights all the ways that the Mosaic tradition offended Gnostic sensibilities. Both Christians and Jews are enamored of the material world. Gnostics, not so much.
0005 The Judeo-Christians believe in one God. So do the Gnostics. But, the one God of the Judeo-Christians is not the same as the Gnostic Source. The Source is a Monad, a complex singularity capable of putting everything into perspective. The Monad is the source of meaning.
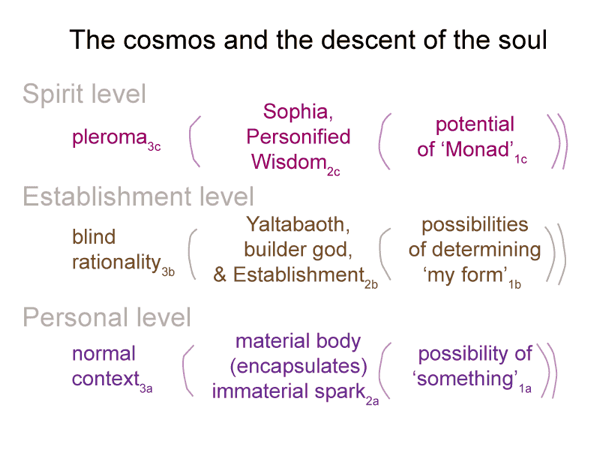
0006 The Monad plays a key role in the Gnostic origin myth. The myth begins with the Source, beyond spoken language and in eternal time. From the Monad, male and female pairs emanated, each according to its aeon. The totality of theaeons is called the “pleroma”.
For the last era, our own, the female of the pair acted without her male consort. He was missing in action, so to speak. Sophia became pregnant with Yaltabaoth, half lion and half snake. She thought to herself, “This is not me.” So she took action, ripping the monster from her womb and hiding him in a cloud.
0007 Born yet aborted, living in a fog, Yaltabaoth decided to build his own world. He decided to create. What else was this demiurge supposed to do? He constructed his own world, which is our universe. He assigned archons (powers and governors) to rule his cosmos.
He was proud of his accomplishments.
Here I make an admission, there is more than one way to spell Yaltabaoth. That exercise is left to the reader.
Okay, back to the story.
0008 What else does this builder god do?
He manufactures our physical bodies. He steals a spark from the Monad (a soul) and pulls it into his world. As the soul descends, Yaltabaoth adds matter. At birth, each person is a spark wrapped in a mortal cloak. The spark is not material. The body is material. The two will war with one another.
0009 Therefore, Sophia pities us and offers this myth. The Self has a mission: Return to the Source. Sophia offers secret knowledge, gnosis, opening a path out of Yaltabaoth’s apparently all-encompassing cloud.
In fact, this myth, which joins cosmic and personal origins, contains a secret. It describes a location where you can find your Self.
0010 Clearly, this origin myth provides meaning. This meaning coincides with the model of social construction in Razie Mah’s How to Define the Word “Religion”. This model is further detailed in A Primer on the Category-Based Nested Form and A Primer on Sensible and Social Construction.
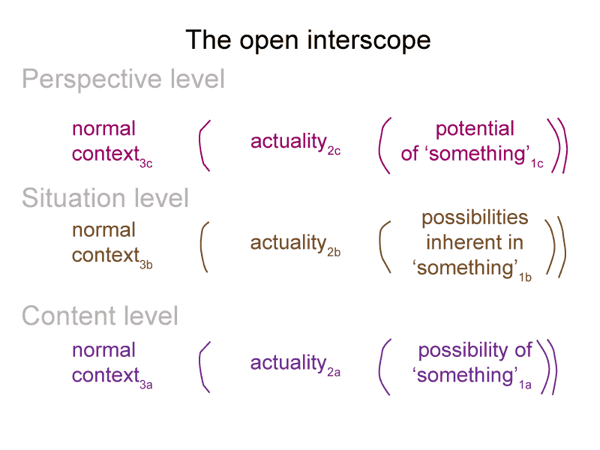
0011 The model is an interscope, a nested form of category-based nested forms. The interscope offers nine slots. Features in the above myth associate with different slots.
Here is the open interscope, before any associations.
0012 My intent is to associate elements of the Gnostic origin myth with slots in the interscope of social construction.
0013 My first association concerns the Monad. The Source has the potential to put the situation into perspective, so it goes into slot 1c. The pleroma automatically becomes the normal context of the perspective level (slot 3c). The pleroma3c brings the possibilities inherent in the Monad1c into relation with the actuality of Sophia2c.
0014 So what is the secret about Sophia?
Sophia is Personified Wisdom. It is not much of a secret. After all, philosophy is rooted in love (philos) and Wisdom (sophia).
0015 If Sophia2c is the mother of the builder god, Yaltabaoth, then I should put the builder god into slot 2b. Yaltabaoth2b situates our universe within the normal context of blind rationality (slot 3b).
Or should I say, “mindless rationality”?
The builder god lives in a cloud, isolated, without awareness of even his own mother. Sure, he snatches sparks from the Monad. He does this blindly. Who knows what spark goes into what body? Yaltabaoth operates in a fog. His archons are like bureaucrats, always following their rulebooks. His social norms? They constitute the Establishment.
The Establishment is the social arm of Yaltabaoth’s evil material world of blind chance and mindless necessity. For teenage Gnostics, that includes all parents. For big-government Gnostics, this includes everything associated with the American Founding.
0016 So, why does the builder god need a spark from the Source in order to animate the human body?
Yaltabaoth is so clueless. He does not realize that the spark comes from the Monad. It is just another thing at his disposal. Obviously, there is no better way to animate humans. He snatches an unwitting soul from its contemplation of Sophia and uses it to bring breath to a breathless body. Inspire. Hold it. Expire. I can feel it.
0017 Here, let me put my associations into the open interscope.